Origen y evolución de la moral
Heraclio Corrales Pavía
Universidad de Málaga
Resumen: Los resultados de la biología evolutiva a menudo son relevantes en ética. Podemos señalar el efecto de esta ciencia en al menos tres niveles de la discusión en ética: el nivel descriptivo, el normativo y el metaético. El objetivo del presente trabajo es mostrar cómo interactúan estas dos disciplinas y cómo es posible justificar el origen de la conducta moral desde el punto de vista de la biología evolutiva. Para ello, será necesario indagar sobre la relación entre genes y entorno, afrontar el reto que supone la falacia naturalista y mostrar las relaciones que se establecen entre las diferentes facultades obtenidas de la evolución.
Palabras clave: Ética, biología evolutiva, sentimientos morales, genes, entorno, cultura, domesticación, altruismo, falacia naturalista, teoría de juegos, estrategia.
Origin and evolution of morality
Abstract: Results from evolutionary biology are sometimes relevant in ethics. It is possible to identify the effects of this science in at least three levels of ethics: the descriptive, the normative and the metaethical one. Our objective is to show what is the interrelation between these two disciplines and how is it possible to justify the origin of the moral behavior from the point of view of evolutionary biology. For doing so, it will be necessary to show the interactions between genes and environment, to face the challenge of the naturalistic fallacy and to show the relationships between the different capacities resulting from the evolutionary process.
Key words: Ethics, evolutionary biology, moral feelings, genes, environment, culture, domestication, altruism, naturalistic fallacy, games theory, strategy.
Recibido: 18/05/2023
Aceptado: 04/10/2023
DOI: 10.24310/nyl.18.2024.16825
1. Introducción
El objetivo del presente trabajo es mostrar qué relación existe entre la biología evolutiva y la ética, estableciendo qué consecuencias se pueden extraer de nuestro conocimiento sobre la evolución de las especies, a efectos de comprender aquellos aspectos de la conducta que serían susceptibles de valoración moral y sobre el modo mismo en el que evaluamos moralmente nuestras acciones. Para ello, comenzaremos con una sección dedicada a establecer unas aclaraciones conceptuales previas, tales como (i) la forma de plantear el papel que juegan el entorno y los genes en la conducta; (ii) en qué aspectos de la ética puede incidir la teoría de la evolución orgánica; (iii) qué consecuencias presenta la aceptación de la tesis que proponemos; y (iv) cuáles serían los rivales intelectuales más importantes de nuestra propuesta. En segundo lugar, analizaremos la acusación de falacia naturalista a nuestra propuesta, mostrando que es posible el desarrollo de una ética naturalista incluso teniéndola en cuenta. A continuación, indagaremos sobre el origen de la conducta moral, para lo que utilizaremos las nociones de estrategia evolutiva estable (EEE) en el contexto de la teoría de juegos y el altruismo, estableciendo los tipos de altruismo que existen y sus relaciones. Luego, señalaremos cómo se relacionan los sentimientos morales con la razón en el juicio moral. Por último, acudiremos al fenómeno de la autodomesticación como proceso evolutivo conducente a la conducta moral y como prueba a favor de la evolución en el tiempo del comportamiento moral.
2. Prolegómenos
A menudo, la cuestión sobre el origen de ciertas conductas se presentas a través de la oposición entre naturaleza y cultura. Este marco aborda las cuestiones sobre el origen de la conducta como fruto de la oposición entre lo innato y lo aprendido, de modo que ciertas conductas estarían codificadas en el ADN y surgirían de forma espontánea, mientras que otras se adquirirían directamente por influencia del entorno cultural. Este enfoque, sin embargo, no nos parece del todo preciso: la inmensa mayoría de las características del ser humano, como ocurre en el resto de los organismos, son el fruto de una relación entre los genes y su entorno, por lo que todo lo que hacemos se puede describir como el resultado de una interacción entre ambos factores. Así, la unidad que produce el fenotipo es la resultante de la expresión de los genes, modulada por el entorno en el que se expresan, entendiendo aquí por entorno todo lo que rodea a los genes, desde las particularidades ambientales concretas hasta el resto de los genes del organismo. Por tanto, lo que se conoce como una conducta innata, como es, paradigmáticamente, la succión en los lactantes, solo se ejercita allí donde el individuo tiene acceso al entorno espaciotemporal en el que se activa dicha conducta; del mismo modo, aprender a usar un arco o un idioma solo es posible si se dan, al mismo tiempo, los requisitos innatos para el aprendizaje de tal conducta y el entorno específico en el que tal desarrollo es posible (Loehlin, 1992). Así, el ser humano, cuyo desarrollo ontogenético se da en un entorno eminentemente social, no solo aprenderá ciertas habilidades en el seno de la sociedad concreta en la que se encuentra inmerso, sino que, además, el modo en el que se desarrollará su inteligencia, su sensibilidad, etcétera, no vendrá enteramente determinado por esto último, sino que también jugarán un papel decisivo en el proceso de aprendizaje los genes que porte dicho individuo (Lewontin, et al., 2017: 276). Por supuesto, no todos los mecanismos requieren de un entorno igual de específico, como queda claro en el ejemplo de la lactancia y de la caza, sino que se sitúan en un continuo de grados de flexibilidad fenotípica, siendo esta «la cantidad de cambios que se produce en un fenotipo […] como respuesta a las alteraciones del ambiente», ambiente que, por cierto, también está sujeto a múltiples cambios (Wilson, 2020: 21). Esto es lo que se conoce como el grado de variación ecofenotípica de los caracteres, que difiere entre los distintos genes (por ejemplo, aquellos que determinan los grupos sanguíneos son de los pocos que se expresan con total independencia del entorno de los individuos).
Así, decir que la conducta en general tiene una base biológica no es incompatible con afirmar que el desarrollo de tal-y-tal práctica o capacidad es al mismo tiempo cultural: hay que buscar las causas de las prácticas en la acción conjunta de ambos factores. Surge inmediatamente la cuestión de si la razón es irrelevante para la conducta: en efecto, parece que en lo dicho esta queda excluida, pero no debemos dejarnos llevar por la primera impresión: la razón es una capacidad cognitiva clave en el pensamiento moral (como mostraremos más adelante), pero es una facultad que tiene su origen en el proceso evolutivo. Así, haya o no sido una adaptación específica para su aplicación en la relación intersubjetiva, tiene el mismo origen evolutivo y ontogenético que el resto de las consideraciones implicadas en la conducta y la deliberación moral, por lo que, por extenso que sea su papel, no es un contraargumento para la tesis del origen biológico de la conducta moral (FritzPatrick, 2021).
Las tesis evolucionistas pueden influir en la ética en, al menos, tres niveles. En primer lugar, en el nivel descriptivo: mostrando cómo es posible la ética desde un punto de vista evolucionista; en segundo lugar, en el nivel normativo: mostrando la pertinencia de propuestas éticas a la luz de las conclusiones de la biología evolucionista, y, por último, en el nivel metaético: revisando propuestas metaéticas a la luz de la biología evolucionista. Por supuesto, no se puede dar el salto sin más del primer nivel al segundo, pero sí es necesario un desarrollo metaético para saber cómo se ven afectadas las «verdades morales» por la biología y qué estatus ontológico, epistémico y lingüístico tienen las «verdades éticas» a la luz de estos descubrimientos (FritzPatrick, 2021) No tratamos tampoco de extraer el contenido de nuestras creencias morales directamente de las tesis evolucionistas, más bien nos parece que, dadas ciertas estructuras cognitivas, existe una «cierta disposición a formar algunos conceptos en lugar de otros» (Diéguez Lucena, 2012: 298). Así, sería del todo sorprendente que seres con compasión, empatía, culpa y vergüenza concluyesen que es una obligación moral torturar a niños pequeños[1], aunque, por supuesto, existen excepciones, por ejemplo, el grado de tolerancia ante las conductas homosexuales difiere extraordinariamente entre las culturas. Sin embargo, como mostraremos en el siguiente apartado, es posible matizar algunas tesis normativas a la luz de la evidencia descriptiva.
Creemos necesario hacer dos advertencias: en primer lugar, que ciertas tendencias (como la de favorecer al propio grupo) sean adaptativas en el sentido que veremos a continuación, no significa que sean buenas moralmente. En el caso de que Sarah Blaffer Hrdy tenga razón y existan bases biológicas para el asesinato de hijastros (como hacen también otros animales) (Cfr. Hrdy, 1979)[2], no se puede concluir de aquí que esa práctica tenga algo de bueno o que esté justificada de algún modo. En segundo lugar, que la conducta moral tenga sus raíces en nuestra configuración genética no significa, al menos en principio, que el progreso que podamos hacer tenga un límite dictado por los genes. Más bien parece que, conociendo los mecanismos concretos que están implicados, podremos crear el entorno específico que genere el desarrollo óptimo de la conducta (Thornill & Palmer, 2000: 179-83). De hecho, en este caso, el conocimiento es poder para alcanzar una sociedad más deseable en términos morales.
Por último, debemos señalar al menos dos rivales de esta teoría. En primer lugar, encontramos la propuesta que hunde las raíces de la moral en la religión. Según sus defensores, el bien y el mal vienen determinados a través de la revelación y las Sagradas Escrituras, que serían la fuente última a la que debemos acudir para saber qué es correcto y qué no. Recuperemos el dilema de Platón: o bien Dios tiene un criterio independiente para establecer qué está bien y qué está mal, por lo que la palabra de Dios es redundante y podemos conocer el bien acudiendo a este criterio, o bien Dios no tiene ningún otro criterio que Su voluntad, por lo que lo que dice es arbitrario y no habría que hacerle demasiado caso (Kitcher, 2011: 166). Por supuesto, esto no debe entenderse como un argumento contra la existencia de Dios mismo, sino sobre el modo en que debemos tratar la ética. En segundo lugar, encontramos a los teóricos de la tabla rasa: aquellos que sostienen «la idea de que la mente humana carece de una estructura inherente y que la sociedad y nosotros mismos podemos escribir en ella a voluntad» (Pinker, 2018: 25). Esta tesis, sin embargo, implica la aceptación de que hubo un momento de la evolución en la que uno grupo de individuos sin ninguna adaptación ni mecanismo cognitivo específico superó a aquellos que llevaban consigo estructuras mentales evolucionadas desde miles a millones de años atrás, lo que nos parece, cuanto menos, implausible (Thornill & Palmer, 2000: 128-9).
3. ¿Falacia naturalista?
Una de las acusaciones más frecuentes que se realizan hacia este planteamiento es que no respeta la separación entre niveles empíricos y normativos. Podemos distinguir al menos cuatro tipos de falacia naturalista: (i) la señalada por Hume, en la que se subraya la diferencia entre el ser y el deber ser; (ii) la falsa equivalencia entre propiedades deseables y éticas, desarrollada por Moore; (iii) la señalada por Frege, Ayer y el primer Wittgenstein, que muestra la imposibilidad de derivar el deber ser del ser; y (iv) el paso de enunciados descriptivos a normativos (Dussel, 2001: 65). No pretendemos negar aquí que las advertencias señaladas deben dejarse de lado para un desarrollo evolucionista de la moral. Tal cosa se ha pretendido en la historia de la mano de los darwinistas sociales, que veían un carácter normativo en las tesis evolucionistas y que proponían, por tanto, un incremento de la lucha social para beneficio de la especie. Esto es exactamente lo que denunciaba Hume (Kitcher, 2011: 255). Sin embargo, es posible señalar modos en los que los descubrimientos empíricos influyen sobre nuestras creencias morales.
En primer lugar, hay que decir que el debate moral no es una esfera de discusión totalmente aislada del plano factual y en la que las tesis del segundo no pueden influir nunca sobre las del primero. El debate sobre cuestiones como los derechos de la mujer o la abolición de la esclavitud no se basa sencillamente en decir que ciertas cosas están bien y que otras están mal, sino que centra la cuestión en aquellos aspectos del mundo que requieren prestar atención a la conducta. Así, cuando Wollstonecraft escribe que las mujeres deben ser mejores madres y esposas (dado por sentado en el siglo XIX) y que las mujeres educadas mejoran también en este aspecto, y, por tanto, sería éticamente correcto dotar a las mujeres de educación (Kitcher, 2011: 257), está diciendo que (i) el que las mujeres sean mejores madres y esposas es moralmente deseable; (ii) la educación es un medio para lograr ese fin moralmente deseable; por consiguiente (iii) luchar por la educación para las mujeres es moralmente deseable.
Así, comprender las causas biológicas de las conductas moralmente reprobables puede llevarnos a tesis normativas que corrijan esas conductas. Digamos, por ejemplo, que: (i) la conducta violenta no es (en general) buena; (ii) el «gen de la violencia», MAO-A, que codifica la enzima monoamina oxidasa A, en su versión de baja actividad parece generar conductas antisociales y violentas en ciertos entornos (Palmqvist Barrena & Andrés Fernández, 2020); (iii) luego debemos comprender qué entornos previenen tales conductas y perseguirlos activamente.
Pero existe un plano más en los que las descripciones del mundo pueden llevarnos a revisar algunas de nuestras tesis normativas. De este modo, podemos establecer qué criterios debemos seguir para evaluar moralmente una acción y qué juicios morales no deberían ser tomados en cuenta. Verbigracia: (i) el grupo al que pertenezco me parece mejor que otros grupos en muchos aspectos; (ii) existe un sesgo cognitivo en el ser humano que nos lleva a favorecer y a tener en mayor estima a aquellos que pertenecen a nuestro grupo (establecido a menudo por criterios más o menos arbitrarios) (Haidt, 2012: 210 y ss.); luego (iii) sabiendo que utilizamos un doble rasero para juzgar a los nuestros y a los demás, no debería tomar como definitivas esas conclusiones, sino que se necesita un criterio independiente para valorar. Así, un conocimiento ampliado del origen de nuestra conducta moral puede llevarnos a entender los límites de nuestros juicios y a comprender qué acción es correcta a la luz de lo que se espera dados ciertos mecanismos.
4. El origen de la conducta altruista
A primera vista, puede parecer sorprendente que, de la selección natural, proceso por el que solo una parte de los organismos vivientes sobreviven dejando descendencia, surjan los mecanismos que propician conductas en las que el beneficiario de la acción no es el propio organismo sino un congénere (normalmente) que, potencialmente, es también un competidor por tales recursos. Sin embargo, han surgido al menos tres líneas argumentativas diferentes (si bien no excluyentes) para dar cuenta de este fenómeno, que aparece en una gran cantidad de especies. Estas propuestas son la selección de parentesco, los beneficios de la cooperación grupal y la selección de grupos (FritzPatrick, 2021: 7). La selección de grupos, defendida entre otros por el ornitólogo Vero C. Wynne-Edwards, señala que la unidad de selección es el grupo, es decir, que la selección natural opera sobre conjuntos de individuos de una misma especie. Así, un grupo que no presente cooperación no tiene las mismas posibilidades de sobrevivir que uno que forme un equipo perfecto. Este mecanismo es supuesto por Darwin a la hora de explicar cómo es posible que las abejas obreras sacrifiquen su vida para defender a la colonia del intruso: una colonia de abejas egoístas que permitiesen que la colonia se hunda antes que sacrificarse ellas parece tener menos posibilidades de sobrevivir que una en la que lo hagan (Diéguez Lucena, 2012: 233). Algunos autores (como Wilson y Sober) han defendido que la selección de parentesco es un caso especial de la selección de grupos (ibidem: 235). Sin embargo, la mayoría de los biólogos han señalado que el mecanismo que se produce aquí es el de la selección de genes o buena parte del conjunto del fenotipo, por lo que quizás es más preciso entender la selección de parentesco como una instancia de la selección de genes (Dawkins, 2017: 138-9)[3]. Dentro de esta propuesta se ha defendido, incluso, que las relaciones afectivas son una exaptación de la conducta de cría llevada a cabo por las madres. Esta visión es consistente, además, con la mayor empatía que presentan comparativamente los animales hembra en gran parte del reino animal, incluida nuestra especie, como muestran algunos experimentos relacionados con las preferencias sociales de las niñas en estadio prelingüístico en contraposición con otros intereses más comunes en los varones (de Waal, 2016: 60).
Sin embargo, nos parece más prometedora la vía del beneficio que otorga la cooperación al miembro altruista del grupo. Utilicemos, para este propósito, el paradigmático juego del dilema del prisionero. Se trata de un escenario hipotético en el que hay dos jugadores cuyas decisiones afectan a su propio beneficio y al del otro jugador, y debiendo decidir, sin saber qué va a hacer el otro, qué opción poner en práctica. Supongamos que dos cómplices de un delito están siendo interrogados en habitaciones diferentes y se les plantea, a ambos, dos opciones: o bien confiesan el delito y aportan la prueba, defraudando al cómplice, o bien ocultan la información en beneficio del cómplice. Los años de cárcel a los que se les condenará a cada uno dependerá del escenario elegido en el modo que se muestra el siguiente cuadro:
|
|
B no aporta la prueba (coopera) |
B aporta la prueba (defrauda) |
|
A no aporta la prueba (coopera) |
1 año, 1 año |
3 años, 0 años |
|
A aporta la prueba (defrauda) |
0 años, 3 años |
2 años, 2 años |
Siendo el número situado en primer lugar el número de años que le corresponden a A y, en segundo, el número de años que le corresponden a B. Tras una breve reflexión, llegaremos a la conclusión de que a ambos delincuentes les interesa defraudar con independencia de lo que haga el otro, más si sabemos que el otro es racional y va a actuar exactamente igual que nosotros (Poundstone, 2018: 204-5). Haga lo que haga el otro, siempre conviene defraudar: si B coopera, entonces, si A coopera, va a la cárcel un año…, pero si, en ese mismo caso, defrauda, entonces se libra de la cárcel. Si B defrauda, entonces, si A coopera, va tres años a la cárcel, pero, si defrauda, son dos años en lugar de tres. Parece que en todo caso lo más conveniente es defraudar.
Esto se puede aplicar a cierta especie de ave en la que, dentro de la bandada, los individuos se desparasitan mutuamente. Un ave no puede quitarse a sí misma los parásitos de la cabeza, pero un congénere sí puede hacerlo por ella. ¿Debería cooperar? Supongamos que un miembro del grupo gasta sus energías en desparasitar a nuestra ave; estamos en el mismo caso que los prisioneros del ejemplo anterior: el máximo beneficio se obtiene no devolviendo el favor y gastando esas energías en el propio interés. Pero estamos suponiendo que el juego se lleva a cabo una sola vez; cuando la interacción es reiterada en el tiempo, las cosas cambian sustancialmente (Dawkins, 2017: 258 y ss.).
En efecto, la situación en la que nos encontramos es subóptima, pero no hay ningún motivo para cooperar incluso así. La cosa cambia, sin embargo, si la competición es reiterada. Si jugásemos varias veces, podríamos obtener un beneficio mayor cooperando de vez en cuando, con la esperanza de que eso motive al otro jugador a cooperar y alcanzar así un beneficio conjunto mayor. Nótese que el número de veces que se juega debe ser indefinido, ya que, en caso contrario, en la última ronda no habrá motivos para cooperar y la situación será similar a la descrita en primer lugar; pero si se sabe que, independientemente de lo que se haga, la última vez no se va a cooperar, tampoco habrá motivos para cooperar la penúltima vez y así sucesivamente. Pues bien, Axelrod organizó un torneo en este escenario entre programas de ordenador, cada uno con una estrategia diferente. Tenían que jugar con el resto de los participantes, con una copia de ellos mismos y con un programa de actuaba aleatoriamente. El vencedor fue un programa muy simple, bautizado como «Donde las dan, las toman» (Tit for Tat), un programa de ordenador que la primera ronda cooperaba y la segunda imitaba la decisión previa del otro jugador. El torneo fue repetido y «Donde las dan, las toman» se alzó de nuevo con la victoria (nótese que el resto de los programadores, amateurs y profesionales, conocían el resultado del primer torneo) (Axelrod, 1984: 27 y ss.).
De esto se sigue que existen estrategias altruistas que superan a las egoístas en juegos planteados de este modo, por lo que, siendo en este caso más beneficioso cooperar, es posible que, en un entorno social, sea adaptativa la evolución de algunos mecanismos de cooperación por el propio beneficio del individuo. No estamos diciendo, por supuesto, que esto implique la conciencia del ser que se comporta así, es suficiente con que existan una serie de estructuras psicológicas que hagan al individuo actuar del modo descrito de modo incluso totalmente programado. Sin embargo, esto no es suficiente, ya que, tratándose de escenarios de tipo estratégico, un mismo modo de actuar puede ser efectivo o no dependiendo de las decisiones de los demás. Así, definamos estrategia como una política preprogramada de comportamiento, del tipo «atacar al adversario; si huye, perseguirlo; si contraataca huir»: el animal se comporta del modo dictado por su estrategia a través de una serie de mecanismos[4]. Una noción clave es la de estrategia evolutivamente estable (EEE): una EEE es una estrategia que, en caso de que se haga mayoritaria en una población, no puede ser superada por ninguna alternativa (Dawkins, 2017: 98-9). Consideremos la estrategia «cooperar de modo incondicionado». Es fácil ver que no se trata de una EEE, ya que los miembros de una sociedad absoluta e incondicionalmente altruista están indefensos ante la aparición de un gorrón (que defraude incondicionalmente), el cual obtendrá una puntuación mucho mayor, por lo que aquellos que siguen su estrategia aumentarán en número[5]. Supóngase que hemos llegado a una composición del grupo en la que se es mayoritariamente gorrón: una estrategia como «Donde las dan, las toman» no tendrá mucho éxito aquí, dado que, antes de dejar de cooperar con nadie, habrá cooperado bastante con individuos que lo han defraudado. Pero supóngase que los miembros de su familia (con los mismos genes) sobreviven algunas generaciones: tendremos un subgrupo que coopera entre sí, con cierta ventaja sobre el resto del grupo. Así, «Donde las dan las toman es una EEE», ya que, siendo mayoritaria, no existe ninguna alternativa que pueda hacerle frente: un tramposo no obtendría el mismo beneficio que el resto y un altruista incondicionado no sería en realidad distinguible (Dawkins, 2017: 260-5). Esta es la idea que subyace en el modelo de Trivers, en el que, a través de una serie de parámetros, se establece la selección del altruismo a partir de la probabilidad de que la ayuda prestada sea devuelta. Estos parámetros son: (i) duración de la vida; (ii) ratio de dispersión; (iii) grado de dependencia mutua; (iv) cuidado parental; y (v) jerarquía de dominancia (donde las relaciones son asimétricas) (Trivers, 1971: 35-57).
Las conclusiones obtenidas hasta este punto son relevantes por dos motivos: en primer lugar, muestran cómo es posible que emerjan tendencias altruistas de la selección natural, esto es, cómo es posible que ciertas conductas de cooperación aumenten la eficacia biológica del individuo y, a la vez, la de otros individuos, no solo la de otros pertenecientes a su grupo familiar; en segundo lugar, muestran que las estrategias deben tener en cuenta las decisiones de los demás; esto es, en caso de que la selección natural haya promovido el altruismo, este no puede ser incondicionado, sino vinculado a las estrategias y comportamientos del resto, lo que nos resulta familiar con el modo en el que vivimos el comportamiento moral. Ahora bien, necesitamos una noción clara de altruismo si queremos establecer que este ha surgido de la evolución y mostrar cómo de un tipo puede seguirse otro.
Podemos distinguir al menos tres nociones de altruismo: el altruismo biológico, el conductual y el psicológico. Entendemos por altruismo biológico un evento causado por un ser vivo, como por ejemplo el babuino macho que se enfrenta al leopardo que amenaza a las crías de su grupo, emparentadas con él, acción que aumenta la eficacia biológica de otro (u otros) organismo(s), las crías en el ejemplo, sin aumentar la suya propia (o incluso disminuyéndola, aunque normalmente en menor medida pues el leopardo suele retirarse ante un ataque concertado de varios machos). Esto no presupone ningún conocimiento sobre los intereses de los demás, ni ningún tipo de intencionalidad por parte del organismo que presenta tal conducta; de hecho, se trata de algo que podemos encontrar incluso en las plantas, que modulan su crecimiento para no dar sombra a sus repollos. El altruismo conductual es aquel en el que el organismo actúa por satisfacer los intereses de otro. Aquí no se entra a considerar las motivaciones que conducen a la conducta altruista: consideramos la acción por sí misma, al margen de que pueda o no estar motivada por fines maquiavélicos (Kitcher, 2011: 18-9). El altruismo psicológico se da si y solo si está movido por el deseo genuino de satisfacer los intereses del otro. Se ha argumentado a veces aquí que el altruismo genuino no existe, ya que, para ayudar a otros cumplir sus fines, es necesario que esto se presente como objeto de un deseo propio, por lo que estaríamos satisfaciendo nuestro propio deseo y, colateralmente, el deseo de los demás (por ejemplo, dar limosna porque es algo que nos reconforta). Es cierto que, para el altruismo psicológico es necesario que los intereses de los otros aparezcan dentro de nuestros propios deseos, pero esa es justamente la diferencia entre un deseo altruista y otro egoísta: el objeto del deseo. Mientras el deseo egoísta tiene por objeto el propio beneficio (materializado en algo en concreto), el deseo altruista tiene por objeto el deseo de otro, y es justamente el hecho de que un deseo pueda tener por objeto los intereses de otro ser vivo lo que debemos explicar (Kitcher, 2011: 19-21).
Así, examinemos el caso de los agentes A y B en los contextos C y C*, siendo C un contexto en el que A conoce los deseos de B y C* un contexto donde A no los conoce. A actúa de modo psicológicamente altruista si y solo si (i) el deseo que ha movido a A en el contexto C es diferente al que hubiese movido a A en el contexto C*; (ii) el deseo de A en C está más cerca de los intereses de B de lo que lo estaría en C*; (iii) el deseo de A en C se basa en la percepción de B en C; y (iv) el deseo de A en C no se basa en ninguna creencia sobre el beneficio personal que se espera de ayudar a B en C. Así, puede entenderse que el altruismo admite grados, medidos por la diferencia entre los deseos de A en C y en C* (Kitcher, 2011: 21-4). Partiendo de la selección del altruismo biológico (si dos individuos aumentan la eficacia biológica del otro más de lo que pueden hacerlo actuando sobre sí mismos, entonces parece que van a tener más eficacia biológica que los demás y, por tanto, tal conducta se va a expandir), es posible que, con el desarrollo de la inteligencia durante la evolución, también se desarrollen capacidades para tener en cuenta la intencionalidad de los demás y prácticas sociales (como el acicalamiento o grooming en los primates) para aumentar la intensidad de las relaciones sociales. Asimismo, aparecen mecanismos de regulación de los recursos (como el sexo, el alimento o el altruismo) y una guía normativa que regula las expectativas y tiene cierto papel coactivo; de este modo, las colonias de monos castigan las trasgresiones de modo violento para preservar el orden social (de Waal, 2016: 236). Una última instancia sería la aparición de instituciones y mecanismos culturales para regular la conducta en nuestros días (FritzPatrick, 2021: 8-10). Así, si la acción de los demás es importante a nivel biológico en un estadio en el que no hay estados mentales (recuperemos el caso de la planta), cabría esperar que, con el desarrollo de la inteligencia, se entrase a considerar de modo consciente la acción de los demás.
5. Razón y emoción en la conducta moral
Se ha dicho hasta ahora que el altruismo puede ser adaptativo y que se puede hablar de altruismo en el plano biológico, en el estrictamente conductual y en el psicológico. Si existe el altruismo psicológico se debe a que es una posibilidad, dadas nuestras estructuras mentales, ya que si no existiese ninguna estructura mental (aunque sea de modo potencial) que permitiese de algún modo que los intereses de los demás aparezcan reflejados en nuestros deseos, esto sencillamente no sería posible. Dado que el altruismo psicológico existe de hecho y que, como se ha mostrado, el altruismo puede tener beneficios para el individuo, examinemos ahora qué mecanismos pueden estar implicados en la generación de este tipo de deseos.
Como decíamos en los prolegómenos, una concepción de la ética de corte kantiano, esto es, que entienda la conducta ética como la que dicta la ley moral que, a su vez, tiene sus orígenes en un uso autónomo de la razón, no es incompatible con nuestro planteamiento. Dado que el uso de la razón es posible gracias a la evolución, deberíamos entender su uso en la ética como un subproducto evolutivo, por lo que estaríamos acudiendo a una facultad fruto de la evolución, si bien no sería una adaptación específica a tales fines, más bien una exaptación[6]. Ahora bien, tenemos motivos para dudar de esto. Un psicópata es una persona que preserva perfectamente la razón, pero carece de sentimientos morales. Dicho de otro modo: un psicópata no es, en lo que a la moral se refiere, un imbécil, sino que, por el contrario, sabe distinguir el bien del mal[7]. Si a un psicópata se le presentasen la teoría del imperativo categórico de Kant, la del espectador imparcial de Adam Smith, la del velo de ignorancia de John Rawls, el principio de utilidad de los utilitaristas o el modo en el que los padres presentan a menudo la cuestión a sus hijos haciendo ver los sentimientos del otro, lo entendería sin problemas; pero respondería que «sencillamente no le importa aumentar la cantidad de felicidad humana del mundo y no le interesa el imperativo categórico, ni nada del resto». Un psicópata sabe lo que está bien y lo que está mal «como un ciego sabe que la hierba es verde», no hay nada en un psicópata que, en principio, impida la comprensión racional de nada de esto, pero no es capaz de encontrar motivos por lo que nada de eso debiera importarle. Recuperemos el caso de un violador de trece años que abusaba de mujeres ciegas porque era más fácil evitar las consecuencias de su acción (Bloom, 2013: 34-5). Por tanto, si existe un caso donde se da la comprensión total de las normas morales desde un punto de vista estrictamente racional y eso no es suficiente para obtener la conducta altruista, debemos concluir que no es, pace Kant, suficiente (aunque sí sea necesaria, como mostraremos más abajo).
Decíamos que aquello de lo que carece un psicópata es el conjunto de los sentimientos morales. De entre todos esos sentimientos se ha destacado la empatía, aunque consideramos pertinente distinguir entre empatía, compasión y ética. Se entiende por empatía la capacidad de ponerse en el lugar del otro. Se cree que las neuronas espejo son las responsables de este fenómeno, si bien es un tema controvertido y no hay una respuesta clara aún sobre el papel de tales neuronas en este proceso. Debemos decir que la empatía no se circunscribe a consideraciones morales, sino que, por el contrario, está presente también en circunstancias moralmente neutras. Cuando bostezamos tras ver a alguien bostezar, es este el mecanismo que está presente, del mismo modo que, según Adam Smith, cuando recomendamos un libro que nos ha gustado o queremos ver nuestra película favorita con otra persona, esperamos sentir por empatía lo que sentimos la primera vez que lo vimos. Se trata de un mecanismo también presente en monos rhesus. La empatía puede ser un mecanismo muy relevante para la moral en el momento en el que el dolor del otro pasa a ser el nuestro propio. La compasión, si bien relacionada a veces, es un mecanismo diferente: se trata del sentimiento de que debemos preocuparnos o cuidar de otro en un momento determinado (Bloom, 2013: 40-3).
Empatía y compasión tienen relevancia moral, pero ni coinciden, ni una implica a la otra. Así, si nos encontramos a un niño ahogándose y sentimos el deber de ayudarlo, obrando en consecuencia, en ningún momento hemos sentido en nosotros mismos la angustia del crío; del mismo modo, si le recomendamos a una amiga el libro que tan interesante nos ha resultado para este trabajo para, como hemos dicho con Adam Smith, sentir por empatía lo mismo que sentimos al leerlo por primera vez, no está obrando aquí ningún tipo de compasión (Bloom, 2013: 44). Frans de Waal sitúa la empatía como el punto de origen de la conducta moral, ya que el reconocimiento de las emociones de los demás a través de este mecanismo genera, por un lado, la comprensión de esas emociones y, de modo derivado, el deseo de intervenir. De Waal señala que se trata de un mecanismo que se encuentra presente en nuestros parientes antropoides, que muestran una clara comprensión de los deseos e intereses de los demás, incluso miembros de otras especies. Así, cita el caso de un macho de bonobo, Kanzi, que comprendía el inglés y «traducía» a su hermana pequeña aquello que se le decía, y el caso de Kuni, una hembra de bonobo que ayudó a un pájaro caído en su jaula a volver a volar y que lo protegió de forma activa de otros miembros del grupo. En ambos casos se dan dos cosas: en primer lugar, una comprensión de los deseos de los demás y, en segundo, el deseo de ayudar (de Waal, 2005: 13-19).
Según de Waal, el origen de la empatía y las relaciones con nuestros familiares y seres queridos aparece como una exaptación del amor maternal. Siendo el vínculo entre una madre y su cría el que, de hecho, caracteriza a los mamíferos, este hecho sería el decisivo a la hora de desarrollar la empatía. Esto se hace más probable si se tiene en cuenta que el primer signo de empatía que se ve en los bebés, llorar cuando escuchan a otro bebé llorar, es más frecuente en niñas que en niños (de Waal, 2005:18). Quizás sea posible argumentar que del hecho de que las mujeres sean más empáticas (sin influencia de la cultura) que los hombres no se sigue que las relaciones de las mujeres son las que están en el origen mismo de la empatía, pues es posible que, dada la empatía, se ha seleccionado una mayor sensibilidad de las mujeres (y de las hembras de los antropoides en general) a fin de una mayor eficacia en la maternidad y en el cuidado de enfermos, ancianos, etc., pues ellas se ocupan (además de recolectar alimentos vegetales en el entorno del campamento) de los individuos desvalidos en las sociedades de cazadores-recolectores mientras los hombres se van a cazar a largas distancias, esto explica también sus mayores competencias lingüísticas, pues los cazadores no deben estar charlando continuamente por el riesgo de espantar las presas. Pero sí resulta muy interesante una de las intuiciones que están detrás de esta propuesta: que el origen de la conducta moral debe de estar en relación con los miembros del grupo más próximos a nosotros.
Una de las cosas que caracteriza nuestra empatía es que es condicional y parcial. Es condicional porque existen casos donde se ve atenuada o exacerbada: así, si está acompañada de la creencia de que el que está sufriendo ha hecho algo malo por lo que se lo merece, el sentimiento es menos intenso, del mismo modo que la fuerza de la emoción aumenta si creemos que es inocente o moralmente bueno (Bloom, 2013: 44). Es parcial porque, dada la misma historia en la que el que sufre es un hombre o un animal, tendemos a sentir mayor rechazo por el caso del hombre (de Waal, 2016: 189) y tiende a ser más fuerte, además de ir acompañada de valoraciones de tipo moral e intelectual positivas, si el que causa empatía es alguien que consideramos miembro del grupo (establecido por criterios arbitrarios) (Bloom, 2013: 123). Por tanto, si efectivamente nuestro sentido de la moral depende de nuestros sentimientos morales, es de esperar que haya brechas en el altruismo que manifestamos dependiendo del beneficiario. Este es el motivo por el que se ha propuesto, como un modo de considerar el progreso moral, la expansión de los círculos de concernimiento moral: así, desde la actitud drásticamente tribal de la hostilidad total hacia las tribus vecinas (Diamond, 2012: 50-2), una de las dimensiones del progreso moral es entender que es necesario dispensar un trato adecuado a todos los miembros de nuestra especie (e incluso de otras, como los grandes simios, los delfines y los elefantes, todas ellas muy encefalizadas) y no solo al resto de los integrantes del grupo (Kitcher, 2011: 302-8).
Por supuesto, nuestro abanico de sentimientos morales es mucho más amplio. Hasta ahora se ha descrito la empatía y la compasión, pero existen aún mecanismos que nos previenen de actuar de modo incorrecto, tales como la culpa y la vergüenza, que pueden entenderse como un castigo autoimpuesto de modo involuntario, relacionado con la creencia de haber actuado de un modo moralmente reprobable, con la particularidad, en el caso de la vergüenza, de que se manifiesta a través del sonrojo en los humanos y que es, por tanto, visible para los demás (Wade, 2014: 58). La culpa y la vergüenza son mecanismos psicológicos que nos previenen de actuar de modo egoísta. Además, tenemos sentimientos morales relacionados con las acciones de los demás, tales como la gratitud, documentada en chimpancés (Cfr. de Waal, 2005: 203), así como el reproche moral (Cfr. de Waal, 2005: 200) y el deseo de castigo al otro (Cfr. Bloom, 2013: 83) o la repugnancia moral, de la que nos ocuparemos en seguida.
Hemos dicho que la razón no es condición suficiente, pero sí necesaria ¿qué razones nos han llevado a decir tal cosa? Principalmente dos: una del campo de la metaética y otra de la ética descriptiva. En primer lugar, si toda nuestra ética se construyese sobre unos sentimientos morales alcanzados a través de la selección natural, no parece posible escapar del relativismo ni del escepticismo: en efecto, se trataría de una ética aplicable sencillamente al entorno en el que nuestra especie ha evolucionado, de modo que, si las abejas tuvieran ética, estaría basada en las relaciones que se dan en el entorno social de las abejas para el entorno ecológico concreto en el que se desarrolla su vida. No podríamos establecer, sin un criterio independiente, un modo con el que conmensurar ambas éticas. Más aún, lo máximo que podríamos decir sobre el bien o sobre el mal es que son normas que han resultado adaptativas y que solo se pueden valorar desde nuestra perspectiva de primate con una serie de sentimientos fijados por la selección natural. Sin embargo, sí existe un criterio independiente, el que nos da la reflexión moral. Es cierto que se ha dicho que esto es posible gracias a la selección natural, pero decir que su uso en ética viene del todo prejuzgado por la evolución sería como decir que la física cuántica venía prejuzgada en la evolución, lo que sería una tesis cuanto menos sorprendente. Si las abejas, además de mecanismos emocionales específicos, tuvieran uso de razón y lenguaje (más allá de sus comunicaciones visuales mediante danzas que indican a sus compañeras la dirección y la distancia a la que se encuentran las flores), quizás sí podríamos establecer un debate moral con ellas (FritzPatrick, 2021: 24-7).
El argumento acerca del papel de la razón que se obtiene en la ética descriptiva es, en cierto modo, parecido. Considere el ejemplo de dos hermanos que, en unas vacaciones, deciden que van a tener una sola relación sexual. Ella toma la píldora anticonceptiva y él, por si acaso, usa preservativo. Después de eso su relación mejora para siempre y no vuelven a repetirlo nunca más. El ejemplo está cuidadosamente diseñado por Haidt para que no sea posible señalar, a primera vista, ninguna consecuencia negativa de ese acto concreto, pero también para que, sin embargo, nos produzca un sentimiento de rechazo a nivel moral. Por supuesto, existen buenas razones para creer que se trata de algo moralmente incorrecto, tales como que es una conducta dañina a nivel psicológico, que existe un grave riesgo de consecuencias graves para la salud de posibles hijos, y que, por esos motivos, bajo ningún concepto debería generalizarse. Pero ese sentimiento de repugnancia es lo que se encuentra en el fondo de la censura moral que tanto tiempo ha pesado sobre los homosexuales. En el caso del incesto y, en general, de las conductas endogámicas, es posible encontrar razones morales y genéticas más allá de nuestros sentimientos de asco para señalar lo incorrecto de la acción; esto no es posible en el caso de la homosexualidad, cuya censura es indefendible a la luz de algunos de los principios más fundamentales obtenidos por la razón (Pinker, 2019: 507-8), por lo que en el primer caso el sentimiento de repugnancia está justificado, mientras que en el segundo debemos concluir que los que lo sentían estaban equivocados al tomar ese sentimiento como autoridad moral. En cualquier caso, la emoción es ciega y debe verse respaldada por una argumentación moral sólida para tener carácter normativo (Bloom, 2013: 131-45 y 154-5).
6. Autodomesticación
Dado que el entorno es el que establece las presiones selectivas por las que un animal evoluciona, es necesario examinar el efecto que ha tenido este en nosotros. Un antropólogo estadounidense, Chris Boehm, ha señalado que, si bien los cazadores-recolectores no tienen un profundo conocimiento de genética, sus prácticas sí que influyen en las disposiciones genéticas de los miembros del grupo. Así, aquellos que quebrantan las normas o muestran una conducta demasiado agresiva, son condenados a muerte o al ostracismo, retirando así sus genes del acervo genético del grupo (de Waal, 2016: 190). Dado que, como hemos señalado, existen algunos genes asociados con la agresión, como el ya citado gen MAO-A (Wade, 2014: 63), (si bien es cierto que la agresión es una conducta compleja que implica muchos genes), parece más probable que los que exhiban este comportamiento censurado en mayor cantidad sean, ceteris paribus los portadores de una mayor cantidad de genes asociados con la violencia, por lo que la tendencia será la supresión de estos genes. Por supuesto, a esto puede objetarse que, en un período de tiempo tan corto, no es posible que la selección tenga efecto. Nos remitiremos aquí al experimento de Dimitri Belyaev, un genetista ruso exiliado por Lisenko a una granja de zorros en Siberia, donde cruzó selectivamente a los zorros más mansos, obteniendo en solo treinta y cinco años zorros tan dóciles como perros. Además, y esto no estaba previsto, la selección de la mansedumbre tuvo efectos pleiotrópicos sobre otros rasgos de los zorros, como la aparición de manchas blancas en el pelaje, colas retorcidas y orejas caídas (Wade, 2014: 171). Más clarificador aún es el experimento que llevó a cabo con ratas, separándolas en dos grupos diferentes, en uno de los casos utilizando como criterio de selección la agresividad y, en el otro, la docilidad. «Tras sesenta generaciones, las ratas mansas se comportaban como mascotas buscadoras de caricias de los cuidadores, mientras que las agresivas parecían enloquecer con solo verlos», mostrando así la explicación genética de ciertos comportamientos (Palmqvist Barrena & Andrés Fernández, 2020).
Podemos definir el síndrome de domesticación como un conjunto universal de rasgos que vienen aparejados a la selección artificial de los animales domésticos, que incluye: (i) cambios en la conducta; (ii) cambios en la composición y el tamaño del cuerpo; (iii) cambios en el tamaño y la composición del cerebro; y (iv) cambios en el color del pelaje (Wright, et al., 2020: 1059). Así, como se muestra en la fig. 1, los animales domésticos presentan una menor proporción entre el logaritmo de la capacidad craneal y el de la masa corporal, es decir, que, manteniendo todo lo demás igual, los animales domésticos han tendido a disminuir su capacidad craneal. Esto implica que la domesticación ha implicado una menor inteligencia en los animales doméstico cuando son comparados con sus versiones salvajes (Palmqvist, et al., 2020: 23). Así, encontramos una relación alométrica negativa entre el tamaño del cuerpo y el cerebro en los animales domésticos durante su desarrollo ontogenético, que, paradójicamente, ha conducido a la hipertrofia de algunos patrones conductuales (Kruska, 1988: 213 y 223). Esto parece desafiante para nuestra tesis, pues, si el ser humano ha sido domesticado y los animales domésticos presentan una reducción del tamaño craneal, ¿no debería haberse producido este fenómeno también en nuestra especie? Sí, y así ha ocurrido: la capacidad craneal se ha reducido sin excepción en todos los animales domesticados, pero en nuestro caso, han existido, además, presiones selectivas a favor del aumento de la encefalización. Las dimensiones del cráneo pueden dividirse en diferentes módulos, y cada uno de ellos con diferentes planos ortogonales. La evolución de los diferentes parámetros es independiente a priori, aunque sí es cierto que existe cierta correlación negativa entre la prominencia de la mandíbula y la cara, y la capacidad cerebral, en torno a un 30%, respecto a sus respectivos agriotipos. Esto implica que la domesticación ha traído como resultado una menor inteligencia en los animales domésticos cuando se comparan con sus versiones salvajes (Palmqvist, et al., 2015: 6 y 8). Así, el aumento de la capacidad craneal del Homo sapiens, junto con el aumento de la sociabilidad con todas sus consecuencias, han conducido a una reducción de capacidad bucal y dientes, con la consecuencia adicional de la neotenia o retención en la madurez de los rasgos de la juventud. Lo que es explicable en parte también a partir de la «expensive tissue» hypothesis, según la cual el excesivo gasto metabólico del cerebro ha implicado de forma paralela la reducción del gasto en otras partes del cuerpo, en particular el tubo digestivo, forzando la evolución hacia una dieta más carnívora (Palmqvist, et al., 2015: 18)[8].
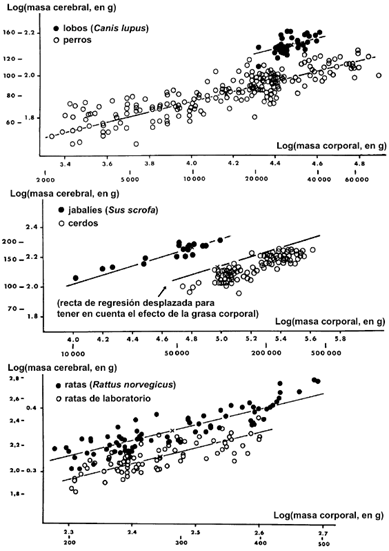
Fig. 1: Relación entre la capacidad cerebral y la masa corporal en animales salvajes y domésticos. Obtenido de (Palmqvist, et al., 2020: 23).
Otra característica que define a los animales domésticos en relación con sus relativos salvajes es la postergación del momento en el que se empieza a interactuar con el entorno. De este modo, el perro (Canis lupus familiaris), comienza a investigar e interactuar con el entorno a las cuatro semanas, cuando ya tienen los órganos sensoriales relativamente desarrollados, y el período de socialización dura cuatro semanas, pudiendo ampliarse mostrando al animal nuevos estímulos. Por el contrario, el período de exploración y socialización de los lobos (Canis lupus) comienza a las dos semanas, cuando tienen desarrollado solo el olfato y dura exclusivamente cuatro semanas. Este es un hecho crucial, ya que la cantidad de estímulos que perciban en las primeras semanas será lo que determinará el modo con el que se relacionarán con esas cosas que aparecen como familiares (Lord, 2012: 113-7). Así, los animales domésticos tienden a explorar e interactuar con el entorno mucho más tarde que los animales salvajes, por lo que quizás el proceso de autodomesticación haya jugado un cierto papel en nuestra ventana de aprendizaje.

7. Conclusión
En el presente trabajo se ha pretendido mostrar qué relaciones guardan la biología evolucionista con la disciplina de la ética. Para ellos, hemos establecido en qué medida y en qué niveles puede afectar la biología evolutiva a los debates en ética y hemos advertido sobre los peligros de caer bajo la llamada falacia naturalista y cómo evitar ese error. A continuación, hemos explicado cómo es posible que del proceso evolutivo surja una conducta altruista y cómo existen buenas razones para creer que el altruismo, en sus diferentes formas, se ha seleccionado naturalmente, hemos seguido entrando a considerar la conducta moral, mostrando que el uso puro de la razón es insuficiente para que se origine dicha conducta moral y acudiendo a los diferentes sentimientos morales de los que da cuenta la psicología moral, estableciendo de este modo qué papel puede tener el legado evolutivo en nuestra especie. Por último, hemos mostrado el proceso por el que es posible que la cultura modifique nuestras bases genéticas en lo relativo a la agresión y la sensibilidad, aludiendo a un proceso con efectos pleiotrópicos que se manifiesta en nuestro fenotipo.
8. Agradecimientos
Agradezco profundamente al profesor Antonio Diéguez Lucena y al profesor Paul Palmqvist Barrena sus directrices y sugerencias de mejoras, así como sus atentos comentarios y matizaciones. Además, agradezco al profesor Palmqvist la cesión de las imágenes y gráficas que se han usado en este artículo.
Bibliografía empleada
Axelrod, R. (1984): The Evolution of Cooperation. Nueva York: Basic Books.
Bloom, P. (2013): Just Babies, the Origins of Good and Evil. New York: Brodway Books.
Dawkins, R. (2017): El gen egoísta extendido. Madrid: Bruño.
de Waal, F. (2005): El mono que llevamos dentro. Barcelona: Tusquests.
de Waal, F. (2016): El bonobo y los diez mandamientos. Barcelona: Tusquests.
Diamond, J. (2012): El mundo hasta ayer. Barcelona: Debate.
Diéguez Lucena, A. (2012): La vida bajo escrutinio. Una introducción a la filosofía de la biología. Barcelona: Biblioteca Buridán.
Dussel, E. (2001): «Algunas reflexiones sobre la ‘falacia naturalista’». Diánoia, 46(46), pp. 65-80. DOI: https://doi.org/10.21898/dia.v46i46.476
FritzPatrick, W. (2021): «Morality and Evolutionary Biology». In: E. N. Zalta, ed. The Standford Enciclopedia of Philosophy. Primavera 2021 ed. Standford: Metaphysics Research Lab, Stanford University. URL: https://plato.stanford.edu/archives/spr2021/entries/morality-biology
Haidt, J. (2012): The righteous mind. Why Good Pepople are Divided by Politics and Religion. Nueva York: Phanteon Books.
Hrdy, S. B. (1979): «Infanticide Among Animals: A Review, Classification, and Examination of the Implications for the Reproductive Strategies of Females». Ethology and Sociobiology, 1(1) pp. 13-40. DOI: https://doi.org/10.1016/0162-3095(79)90004-9
Kitcher, P. (2011): The Ethical Project. Massachusetts: Harverd University Press.
Kruska, D. (1988): «Mammalian Domestication and its Effect on Brain Structure and Behavior». Institut für Haustierkunde, 17 pp. 211-250.
Lewontin, R. C., Rose, S., & Kamin, L. J. (2017): Not in our Genes. Biology, Ideology, and Human nature. Chicago: Haymarkets Books.
Loehlin, J. C. (1992): Genes and environment in personality development. Sage Publications, Inc.
López, A. (2018): «Hormigas bala: el ritual de iniciación más cruel y doloroso del mundo». Cultura colectiva, 30 Enero.
Lord, K. (2012). «A Comparision of the Sensory Development of Wolves and Dogs». International journal of behavioural biology, 119(2), pp. 110-1120. DOI: https://doi.org/10.1111/eth.12044
Palmqvist Barrena, P., & Andrés Fernández, A. V. d. (2020). «Covid-19: ¿nos ha vuelto más o menos solidarios?». The Conversation.
Palmqvist, P., Andrés, V. d. L. M. A., & Olivero, J. (2020). «La ciencia y el animalismo». Encuentros en la biología, 5(141), pp. 19-33.
Palmqvist, P., Pérez-Claros, J. A., & Jiménez-Arenas, J. M. (2015). «Neurocranium versus Face: A Morphometric Approach with Classical Anthropometric Variables for Characterizing Patterns of Cranial Integration in Extant Hominoids and Extinct Hominins». Ploss One, 10(7). DOI: 10.1371/journal.pone.0131055. eCollection
Pinker, S. (2018). La tabla rasa. La negación moderna de la naturaleza humana. Barcelona: Paidós.
Pinker, S. (2019). En defensa de la Ilustración. Barcelona: Paidós.
Poundstone, W. (2018). El dilema del prisionero. Madrid: Alianza.
Thornill, R., & Palmer, C. T. (2000). A Natural History of Rape. Biological Bases of Sexual Coercion. Massachusetts: Massachusetts Institute of Technology.
Trivers, R. L. (1971). «The Evolution of Reciprocal Altruism». The Quarterly Review of Biology, 46(1), pp. 35-57. DOI: 10.1086/406755
Wade, N. (2014). Una herencia incómoda. Genes, razas, e historia humana. Barcelona: Ariel.
Wilson, E. O. (2020). Génesis. El origen de las sociedades. Barcelona: Crítica.
Wright, D., Henriksen, R., & Johnson, M. (2020). «Defining the Domestication Syndrome: Comment on Lord et al. 2020». Cell Press Reviews, 35(12), pp. 1059-1060. DOI: https://doi.org/10.1016/j.tree.2020.08.009
Heraclio Corrales Pavía
heracliocorrales@uma.es